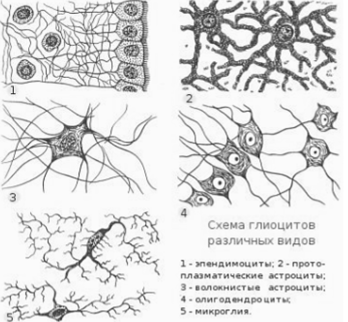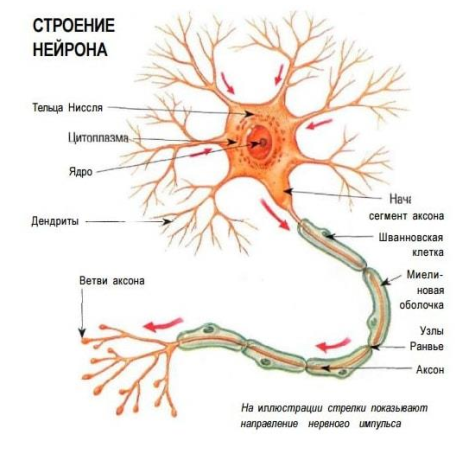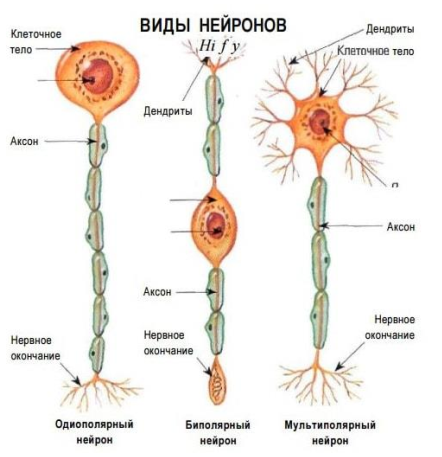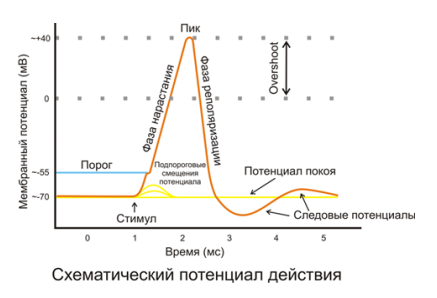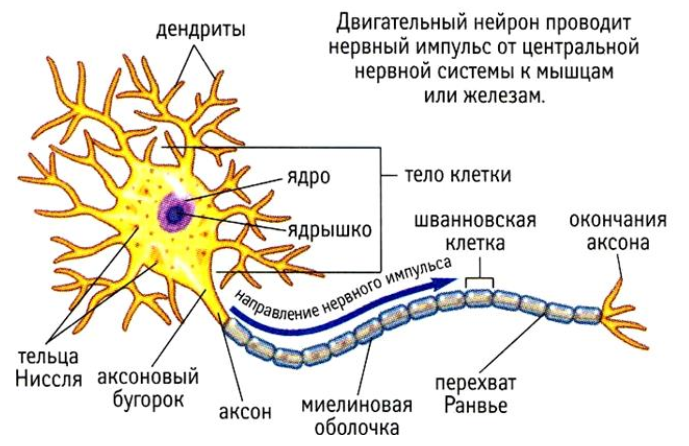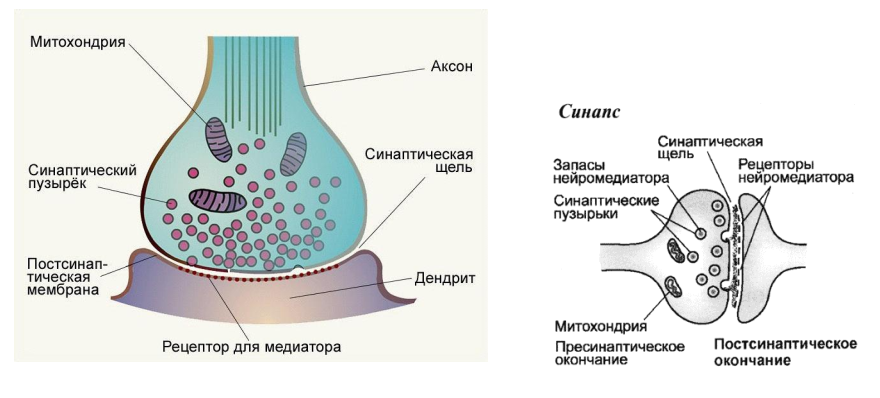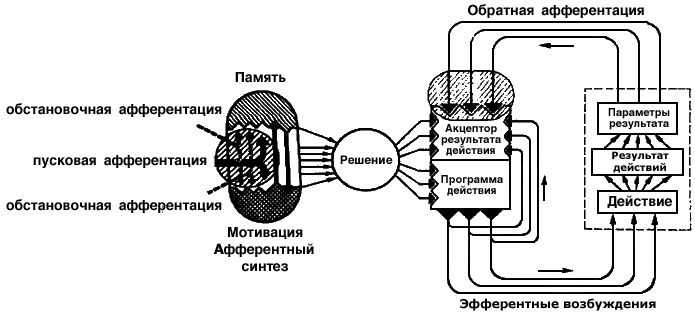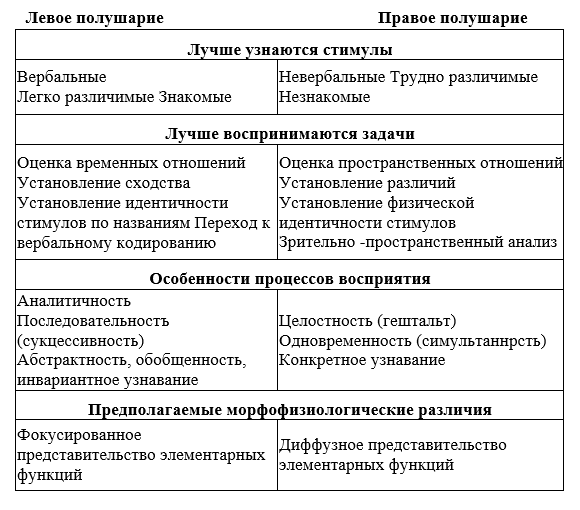
Тема 1. ПРЕДМЕТ И ЗАДАЧИ ПСИХОФИЗИОЛОГИИ
Психофизиология (психологическая физиология) научная дисциплина, возникшая на стыке психологии и физиологии, предметом её изучения являются физиологические основы психической деятельности и поведения человека.
Термин «психофизиология» был предложен в начале XIX века французским философом Николя Массиасом и первоначально использовался для обозначения широкого круга исследований психики, опиравшихся на точные объективные физиологические методы (определение сенсорных порогов, времени реакции и т.д.). Психофизиология - естественно-научная ветвь психологического знания, поэтому необходимо определить её положение по отношению к другим дисциплинам той же ориентации:
• физиологической психологии;
• физиологии высшей нервной деятельности;
• нейропсихологии.Наиболее близка к психофизиологии физиологическая психология - наука, возникшая в конце XIX века как раздел экспериментальной психологии. Термин «физиологическая психология» был введён В. Вундтом для обозначения психологических исследований, заимствующих методы и результаты исследований у физиологии человека. В настоящее время физиологическая психология понимается как отрасль психологической науки, изучающая физиологические механизмы психической деятельности от низших до высших уровней её организации.
Таким образом, задачи психофизиологии и физиологической психологии практически совпадают, и в настоящее время различия между ними носят в основном терминологический характер. Однако был период в истории отечественной психофизиологии, когда терминологические различия были использованы для того, чтобы обозначить продуктивность складывающегося в физиологии функционально-системного подхода к изучению психики и поведения человека. Выделение психофизиологии как самостоятельной дисциплины по отношению к физиологической психологии было проведено А.Р. Лурией. Согласно его представлениям, физиологическая психология изучает основы сложных психических процессов - мотивов и потребностей, ощущений и восприятия, внимания и памяти, сложнейших форм речевых и интеллектуальных актов, т.е. отдельных психических процессов и функций. Она образовалась в результате накопления большого объёма эмпирического материала о функционировании различных физиологических систем организма в разнообразных психических состояниях.
В отличие от физиологической психологии, где предметом является изучение отдельных физиологических функций, предметом психофизиологии служит поведение человека или животного. При этом поведение оказывается независимой переменной, тогда как за-висимой переменной являются физиологические процессы. По Лурии, психофизиология - это физиология целостных форм психической деятельности, она возникла в результате необходимости объяснить психические явления с помощью физиологических процессов, и поэтому в ней сопоставляются сложные формы поведенческих характеристик человека с физиологическими процессами разной степени сложности.
Истоки этих представлений можно найти в трудах Л.С. Выготского, который первым сформулировал необходимость исследовать проблему соотношения психологических и физиологических систем. Теоретико-экспериментальные основы этого направления составляет теория функциональных систем П.К. Анохина (1968), базирующаяся на понимании психических и физиологических процессов как сложнейших функциональных систем, в которых отдельные механизмы объединены общей задачей в целые, совместно действующие комплексы, направленные на достижение полезного приспособительного результата.
Наконец, следует указать на соотношение физиологии ВНД и психофизиологии. Высшая нервная деятельность (ВНД) - понятие, введенное Н.П. Павловым, - в течение многих лет отождествлялось с понятием «психическая деятельность». Таким образом, физиология высшей нервной деятельности представляла собой физиологию психической деятельности, или психофизиологию.
Представим себе мозг живого человека: он выглядит как небольшое овальное тело с неровной поверхностью, состоящее из податливого желеподобного вещества. Каким образом это тело (средний вес которого составляет 1500 г) продуцирует мысли и чувства, управляет тонкими движениями руки художника? Каким образом возникающие в нём процессы связываются с мировой культурой: философией и религией, поэзией и прозой, добротой и ненавистью?
В этих вопросах в предельно заострённой метафорической форме можно выразить суть основной проблемы психофизиологии - проблемы соотношения мозга и психики, психического и физиологического.
История проблемы и варианты решения. Проблема соотношения психики и мозга, души и тела, разведение их по разным уровням бытия имеет глубокие исторические традиции, прежде всего, традиции европейского мышления, существенно отличающегося от многих восточных систем миросозерцания.В европейской традиции термины «душа» и «тело» впервые стал рассматривать с научных позиций выдающийся философ и врач Рене Декарт, живший в XVII в. По Декарту, тело - это автомат, действующий по законам механики и только при наличии внешних стимулов. Именно Декарт выдвинул идею рефлекса как машинообразного ответного поведенческого акта (хотя сам термин «рефлекс» был предложен спустя столетие ). Душа, напротив, особая сущность (субстанция), состоящая из непротяжённых явлений сознания - «мыслей». Именно мысль представляет наиболее доступный объект самонаблюдения. Отсюда знаменитое утверждение: «Я мыслю, следовательно, я существую».
Итак, Декарт рассматривал душу и тело как две самостоятельные, независимые субстанции. Однако как душа может влиять на деятельность тела, так и тело, в свою очередь, способно сообщать душе сведения о внешнем мире. Для объяснения этого взаимодействия Декарт предположил, что в мозгу человека имеется специальный орган — шишковидная железа (эпифи`з)— посредник между душой и телом. Воздействие внешнего мира вначале передается нервной системой, а потом тем или иным способом «некто» (гомункулус) расшифровывает содержащуюся в нервной деятельности информацию.
Таким образом, Декарт, чётко разделив тело и душу человека, впервые поставил проблему их соотношения и дал первый вариант её решения, получивший название психофизического и/или психофизиологического параллелизма. Учение Декарта, исходящее в объяснении сущего из наличия двух противоположных начал — материального и духовного, получило название дуализма Декарта.
Сходных взглядов придерживались многие современники и последователи Декарта, например, выдающийся философ и математик Г. Лейбниц. Согласно его представлениям, душа и тело действуют независимо и автоматически в силу своего внутреннего устройства, но действуют удивительно согласованно и гармонично, подобно паре точных часов, всегда показывающих одно и то же время.
Психофизическая проблема. Как подчёркивает известный отечественный историк психологии М.Г. Ярошевский (1996), Декарт, Лейбниц и другие философы анализировали в основном психофизическую проблему. При её решении речь шла о включении души (сознания, мышления) в общую механику мироздания, о её связи с Богом. Иными словами, для философов, решающих эту проблему, важно было собственно место психического (сознания, мышления) в целостной картине мира.
Психофизиологическая проблема заключается в решении вопроса о соотношении между психическими и нервными процессами в конкретном организме (теле). В такой формулировке она составляет основное содержание предмета психофизиологии. Первое решение этой проблемы можно обозначить как психофизиологический параллелизм. Суть его заключается в противопоставлении независимо существующих психики и мозга (души и тела). В соответствии с этим подходом психика и мозг признаются независимыми явлениями, не связанными между собой причинно-следственными отношениями.
В то же время наряду с параллелизмом сформировались ещё два подхода к решению психофизиологической проблемы:
- психофизиологическая идентичность, которая представляет собой вариант крайнего физиологического редукционизма, при котором психическое, утрачивая свою сущность, полностью отождествляется с физиологическим. Примером такого подхода служит известная метафора: «Мозг вырабатывает мысль, как печень - желчь».
- психофизиологическое взаимодействие, представляющее собой вариант паллиативного, т.е. частичного, решения проблемы. Предполагая, что психическое и фи-зиологическое имеют разные сущности, этот подход допускает определённую степень взаимодействия и взаимовлияния.
Несмотря на многие достижения психофизиологии, особенно в последние десятилетия, психофизиологический параллелизм как система взглядов не отошёл в прошлое. Известно, что выдающиеся физиологи XX в. Шерингтон, Эдриан, Пенфилд, Экклс придерживались дуалистического решения психофизиологической проблемы. Согласно их мнению, при изучении нервной деятельности не надо принимать во внимание психические явления, а мозг можно рассматривать как механизм, деятельность определенных частей которого, в крайнем случае, параллельна разным формам психической деятельности. Целью психофизиологического исследования, согласно их мнению, должно являться выявление за-кономерностей параллельности протекания психических и физиологических процессов.
Взаимосвязь психики и мозга. Многочисленные клинические и экспериментальные данные, накопленные в науке в последние десятилетия, свидетельствуют, однако, что между психикой и мозгом существует тесная и диалектическая взаимосвязь. Воздействуя на мозг, можно изменить и даже уничтожить дух (самосознание) человека, стереть личность, превратив человека в зомби. Сделать это можно следующими способами:
- химически, используя психоделические вещества (в том числе наркотики);
- «электрически» (с помощью вживлённых электродов);
- анатомически, прооперировав мозг.В настоящее время с помощью электрических или химических манипуляций с определёнными участками головного мозга человека изменяют состояния сознания, вызывая различные ощущения, галлюцинации и эмоции.
Всё вышесказанное неопровержимо доказывает прямое подчинение психики внешним физико-химическим воздействиям. Более того, в последнее время всё больше и больше накапливается данных о том, что психологические состояния человека тесно связаны с наличием или отсутствием того или иного химического вещества в мозге.
С другой стороны, всё, что глубоко затрагивает психику, отражается также и на мозге, и на всём организме. Известно, что горе или сильная депрессия могут привести к телесным (психосоматическим) заболеваниям. Гипноз может вызвать различные соматические расстройства или, наоборот, способствовать излечению. Широко известны поразительные эксперименты, которые осуществляют йоги со своим организмом. Более того, такие психокультурные явления, как нарушение «табу» или колдовство, у примитивных народов могут вызвать смерть даже здорового человека. Есть свидетельства, что религиозные чудеса (явления Богоматери, Святых икон и т. п.) способствовали исцелению больных с различной симптоматикой. Интересно в этой связи, что эффект плацебо, т. е. эффект нейтрального вещества, которое применяется вместо «ультрасовременного» лекарства, действенен для одной трети больных, независимо от их социального статуса, культурного уровня, вероисповедания или национальности.
В целом приведённые выше факты однозначно свидетельствуют о том, что столь тесную взаимосвязь между мозгом и психикой нельзя объяснить с позиций физиологическо-го параллелизма. Важно, однако, подчеркнуть и другое. Отношение психики к мозгу нельзя понимать как отношение продукта к производителю, следствия к причине, поскольку продукт (психика) может и часто очень эффективно воздействует на своего производителя - на мозг. Таким образом, между психикой и мозгом, психическим и физиологическим, по-видимому, существует диалектическая, причинно-следственная связь, ещё не получившая полного объяснения.
Современные варианты решения психофизиологической проблемы можно систематизировать следующим образом:
1. Психическое тождественно физиологическому, представляя собой не что иное, как физиологическую деятельность мозга. В настоящее время эта точка зрения формулируется как тождественность психического не любой физиологической деятельности, но только процессам высшей нервной деятельности. В этой логике психическое выступает как особая сторона, свойство физиологических процессов мозга или процессов высшей нервной деятельности.2. Психическое — это особый (высший) класс или вид нервных процессов, обладающий свойствами, не присущими всем остальным процессам в нервной системе, в том числе процессам ВНД. Психическое - это такие особые (психонервные) процессы, которые связаны с отражением объективной реальности и отличаются субъективным компонентом (наличием внутренних образов и их переживанием).
3. Психическое, хотя и обусловлено физиологической (высшей нервной) деятельностью мозга, тем не менее НЕ ТОЖДЕСТВЕННО ей. Психическое несводимо к физиологическому, как идеальное к материальному или как социальное к биологическому.
Ни одно из приведённых решений не получило общего признания, и работа в этом направлении продолжается. Наиболее существенные изменения в логике анализа проблемы «мозг - психика» повлекло за собой внедрение в психофизиологию системного подхода.